Investigadores del Centro de Biotecnología y Genómica Vegetal (CBGP, UPM-INIA), en colaboración con la Universidad de Lleida-Agrotecnio y la Institución Catalana de Investigación y Estudios Avanzados (ICREA), han conseguido producir los primeros cereales transgénicos que expresan dos componentes clave de la nitrogenasa, la enzima que fija el Nitrógeno atmosférico convirtiéndolo en amoníaco.
Cada componente se produjo en una línea de plantas transgénicas distinta y se demostró que es biológicamente activo tanto in vitro como en plantas vivas. Estas plantas transgénicas aún no pueden fijar su propio Nitrógeno porque se necesitan componentes adicionales para reconstruir la enzima nitrogenasa completa, pero el trabajo es innovador porque demuestra por primera vez que es posible expresar estas proteínas altamente sensibles al Oxígeno de manera estable en las plantas, y que las proteínas conserven sus actividades.
Los cultivos requieren Nitrógeno para su crecimiento y productividad porque es un componente importante del ADN, las proteínas, la clorofila y las moléculas de almacenamiento de energía, como el trifosfato de adenosina (ATP). La mayoría de los cultivos dependen del suministro de Nitrato y Amonio a partir de los fertilizantes químicos industriales, pero más de la mitad de estos insumos no se asimilan y se pierden o se infiltran en ríos y lagos como una fuente relevante de contaminación.
Los cultivos de leguminosas como los chícharos y los frijoles albergan bacterias que convierten el gas Nitrógeno directamente en Amoníaco utilizando una enzima llamada nitrogenasa. Este proceso se conoce como fijación biológica de Nitrógeno. La introducción de genes de nitrogenasa en plantas de cultivo les proporcionaría la maquinaria necesaria para fijar el Nitrógeno de forma independiente. Sin embargo, el proceso es extremadamente complejo porque se requieren muchas proteínas individuales diferentes, no sólo como componentes estructurales directos de la nitrogenasa, sino también proteínas accesorias necesarias para su ensamblaje y la provisión de energía. Los principales componentes proteicos también son extremadamente sensibles al Oxígeno.
Los investigadores superaron este cuello de botella crítico al producir dinitrogenasa reductasa funcional (proteína Fe, NifH) y el cofactor de nitrogenasa maturasa (NifB) en líneas de arroz transgénico diferentes. La investigación sobre la expresión de nitrogenasa generalmente se lleva a cabo en plantas modelo de laboratorio. Sin embargo, al centrarse en el arroz, un importante cultivo básico que proporciona la principal o única fuente de calorías para más de 2500 millones de personas en los países en desarrollo, la importancia y el impacto de los resultados de los estudios aumentan sustancialmente.
El investigador principal del proyecto, el Dr. Luis Rubio, dice: «Este es un gran avance de la bioingeniería, ya que derriba dos obstáculos técnicos y muestra el camino para generar cereales fijadores de Nitrógeno». El logro elimina una de las principales limitaciones que obstaculizan la fijación biológica de Nitrógeno en los cultivos y prepara el escenario para el ensamblaje de un complejo de nitrogenasa completo y funcional en las plantas.
El trabajo adicional para establecer plantas que contengan la nitrogenasa completa tendría un impacto duradero en la seguridad alimentaria mundial. El Dr. Paul Christou, profesor de investigación ICREA y líder del proyecto en la Universidad de Lleida-Centro Agrotecnio, dice: «Uno de los principales impactos del trabajo a largo plazo será en los países de ingresos bajos y medios, que no pueden permitirse costosos fertilizantes nitrogenados».
La investigación relacionada ha sido publicada en Communications Biology y ACS Synthetic Biology.
Debajo de la superficie del suel0, las raíces de las plantas se desarrollan continuamente y es por medio de ellas, que las plantas obtienen agua y minerales del suelo. Aprender cómo los diferentes sistemas de raíces afectan el rendimiento de los cultivos ha sido un desafío para los investigadores.
«Sabemos mucho menos sobre las características de las raíces y cómo afectan los rendimientos de los cultivos en comparación con las características de las hojas», dice Maryse Bourgault, investigadora de la Universidad de Saskatchewan, Canadá.
Bourgault es la autora principal de un nuevo estudio en el que se investigan los vínculos entre los sistemas de raíces y el rendimiento en cultivos de lentejas y chícharos en zonas semiáridas. Este estudio fue publicado en “The Plant Phenome Journal”, una publicación de la Crop Science Society of America.
Un gran porcentaje de las exportaciones mundiales de lentejas se originan en las Grandes Llanuras del Norte en los Estados Unidos y Canadá. En estas regiones semiáridas, casi 4.5 millones de hectáreas (superficie mayor de la que le corresponde al estado de Maryland), se utilizan para producir cultivos de chícharos y lentejas.
Bourgault y sus colegas encontraron que las variedades de chícharos y lentejas de mayor rendimiento tenían estructuras de sistemas de raíces bastante diferentes al resto.
En lentejas, sistemas de raíces grandes se correlacionaron bien con altos rendimientos. «Las plantas de lentejas tienden a ser pequeñas, por lo que los mejoradores han estado tratando de hacerlas más grandes y altas», dice Bourgault. «Si estamos presionando para tener plantas de lentejas más grandes, también deberíamos hacer selección para sistemas de raíces de lentejas más grandes».
En chícharos, la situación fue más compleja. Las variedades de chícharos de mayor rendimiento tendían a tener sistemas de raíces de tamaño promedio.
«Creemos que el crecimiento de las raíces en los chícharos puede depender más del momento durante la temporada de crecimiento de las plantas», dice Bourgault. Los investigadores creen que la mayor parte del crecimiento de las raíces debe ocurrir antes de que florezcan las plantas de chícharos. «Una vez que ocurre la floración, toda la energía de la fotosíntesis debe ir al desarrollo de las vainas de chícharos más que al crecimiento de la raíz».
El hallazgo de que diferentes sistemas de raíces conducirían a mayores rendimientos en diferentes cultivos fue un poco contra la corriente. El patrón predominante era que un sistema radicular «profundo pero delgado» sería más benéfico para todos los cultivos en zona semiáridas.
La idea “profundo pero delgado” se basó en la investigación del cultivo de trigo», dice Bourgault. Un estudio encontró que una variedad de trigo con un sistema de raíces delgado pero profundo, se adaptaba mejor a las regiones semiáridas.
Eso pudo ser porque las plantas de trigo con sistemas de raíces profundas tenían acceso al agua que se encontraba a mayor profundidad en el suelo. Sin embargo, la delgadez del sistema de raíces significó que las plantas no habían invertido demasiados recursos en las raíces, lo que permitió tener mayores rendimientos de grano.
«La idea de ‘profundo pero delgado’ ha sido muy popular», dice Bourgault. «Demostramos que ésta no es una verdad universal. Lo que podría ser cierto para el trigo puede no serlo para otros cultivos».
Los hallazgos de Bourgault también resaltan las ventajas de estudiar directamente los sistemas de raíces de los cultivos, especialmente en zonas semiáridas, donde el agua suele ser escasa. «Creo que estamos en el punto en el que las próximas grandes ganancias en los rendimientos en las zonas semiáridas vendrán al contemplar seriamente los sistemas de raíces y comprender cómo funcionan», dice.
El sitio de estudio de la presente investigación estaba ubicado en el Centro de Investigación Agrícola del Norte de la Universidad Estatal de Montana. De 2017 a 2019, los investigadores escanearon y midieron la longitud de las raíces en 29 variedades de chícharos y 25 de lentejas.
«Descubrimos que había muchas diferencias entre las variedades de chícharos y lentejas en lo que se refiere a cuánto invierten en los sistemas de raíces», dice Bourgault.
Bourgault y sus colegas ahora buscan identificar genes o regiones genéticas en lentejas y chícharos asociados con datos sólidos de rasgos de raíces. «Eso podría ser potencialmente un cambio de juego para la introducción de características de la raíz en los programas de mejoramiento», dice ella.
Suelos, diversidad microbiana y función ecosistémica
Los suelos son recursos no renovables y constituyen sistemas complejos por su heterogeneidad (variaciones del suelo por áreas, temporadas y circunstancias). Esta complejidad dificulta la selección de indicadores que sean útiles para medir y monitorear la calidad del suelo y que puedan integrarse a programas de manejo y conservación. Los suelos son la matriz en la que ocurren los procesos biogeoquímicos que permiten el reciclaje y la reincorporación de nutrientes, de un modo que permita el funcionamiento de los ecosistemas naturales y de los sistemas socio ambientales, y que permita, en última instancia, la subsistencia de las poblaciones humanas.
Los microorganismos del suelo desarrollan un papel fundamental en los procesos biogeoquímicos y de esta forma contribuyen a su fertilidad, así como a la productividad y la salud de las plantas. Adicionalmente, estas comunidades son lo suficientemente sensibles a las variaciones ambientales y a los cambios de uso de suelo como para poder fungir como bioindicadores de cambios en su calidad.
Los organismos del suelo están asociados a la diversidad vegetal. La biota del suelo se ve afectada por la cantidad y calidad del material vegetal que se le incorpora durante el proceso de descomposición y por las diferentes condiciones ambientales (incluso a escala microscópica), que generan las comunidades vegetales. Se ha documentado que esta relación es bidireccional: los microorganismos del suelo afectan la calidad del tejido de las plantas, y las plantas a su vez afectan a las comunidades de microorganismos del suelo. Por lo que un mejor entendimiento de estas relaciones promete la posibilidad de identificar aspectos del suelo (físicos y biológicos) que puedan incorporarse como indicadores del estado general del sistema (Suelo-Planta-Microorganismos), en relación con las propiedades ideales del mismo (por ejemplo, resiliencia ante perturbaciones o resistencia a las plagas)
Diversidad Microbiana de Suelos y Funcionamiento de los Ecosistemas
Conocer el componente microbiológico de los suelos de los sistemas productivos es esencial para la predicción y toma de decisiones sobre clima futuro, fertilidad del suelo, remediación de desastres causados por daños industriales, almacenaje de Carbono, reciclaje de nutrientes y bienestar humano; es decir, para que la sostenibilidad de estos Sistemas Socio-Ecológicos (SSE) sea posible. Sin embargo, la investigación sobre la relación entre la diversidad microbiana y su contribución al funcionamiento de los ecosistemas aún es escasa.
En el informe de 2010 para la Línea Base Nacional de Degradación de Tierras y Desertificación, la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT), la Comisión Nacional Forestal (CONAFOR), la Universidad Autónoma de Chihuahua (UNICH) y el Sistema Nacional de Lucha Contra la Desertificación (SINADES), reportaron que el 90% de los suelos del país tiene algún tipo de degradación; es de esperarse que, dado el patrón de cambio de uso de suelo, el porcentaje en la actualidad sea mayor. La acelerada degradación de los suelos y la consecuente pérdida de biodiversidad ponen en grave riesgo el mantenimiento y la productividad de los ecosistemas naturales, los agroecosistemas y los sistemas agroforestales, lo que constituye un grave problema nacional.
En México, la CONAFOR ha iniciado el Sistema Nacional de Monitoreo de Degradación de Tierras y Desertificación, y la SEMARNAT ha desarrollado la Estrategia Nacional de Manejo Sustentable de Tierras. No obstante, estos planes de alcance nacional no toman en cuenta el monitoreo, conservación y aprovechamiento de la diversidad microbiana.
Dado el papel clave de los microorganismos en los procesos que ocurren en los suelos, es necesario considerar el componente microbiano en la elaboración de indicadores de degradación y en cualquier plan de mitigación y restauración sostenible de suelos.
Estudiar la Diversidad Microbiana y las Funciones Ecosistémicas
Si bien aún es escasa, la investigación sobre los mecanismos que relacionan la diversidad de microorganismos con las funciones ecosistémicas que desempeñan, ha cobrado gran ímpetu en la última década. Este tipo de investigaciones son relevantes debido a la necesidad de entender mejor, por ejemplo, las consecuencias del cambio climático, y son necesarias para mejorar las predicciones y decisiones relacionadas con el manejo de ecosistemas. Hoy este tipo de investigación es posible gracias a los avances tecnológicos que permiten el estudio de la diversidad microbiana en ambientes naturales, de manera independiente del cultivo en laboratorio.
Ejemplo de estos avances son, entre otros, las herramientas de tipo genómico. Éstas permiten estudiar a las comunidades microbianas completas, con lo cual ha sido posible indagar lo que sucede cuando se altera la biodiversidad de dichas comunidades, en relación con las funciones que cumplen; a esto se le llama metagenómica. Históricamente, para conocer la diversidad de las comunidades microbianas era necesario trabajarlas de manera controlada en el laboratorio en cajas Petri (en los llamados cultivos selectivos).
No obstante, mediante esta técnica sólo se logra caracterizar alrededor del 1-10 % de los microbios totales de las muestras ambientales, lo cual no es realista si queremos conocer su diversidad en los ecosistemas naturales. Para ello, la alternativa es obtener todo el material genético de las muestras ambientales.
Investigación para Desarrollar Indicadores de Sostenibilidad
La investigación en ciencias de la sostenibilidad tiene como objetivo generar conocimiento útil para la sociedad y para la toma de decisiones que faciliten el tránsito hacia un futuro sostenible. El que éste conocimiento tenga una aplicación útil se denomina vinculación y forma parte de lo que Brandt y colaboradores denominaron investigación transdisciplinaria en su artículo “A Review of Transdisciplinary Research in Sustainability Science”. Este proceso de vinculación se desarrolla en las denominadas entidades de frontera. La función de estas entidades es la coproducción, traducción, comunicación y mediación del conocimiento entre actores académicos y sociales, con la finalidad de que se tomen las mejores decisiones orientadas hacia la sostenibilidad.
Dentro del Instituto de Ecología de la UNAM, el Laboratorio Nacional de Ciencias de la Sostenibilidad (LANCIS) se concibe como una entidad de frontera y por ello se compromete con la vinculación. En este tipo de entidades es muy importante traducir los hallazgos de manera que sean accesibles conceptualmente para cualquier persona, para que así esta información pueda ser considerada en las decisiones pertinentes. De esta forma, el trabajo transdisciplinario en una entidad de frontera debe contar con la participación de académicos de otras áreas de conocimiento más allá de la Ecología, así como de distintos actores sociales, para lograr la traducción, estimación de riesgos, generación de escenarios y desarrollo de plataformas que sostengan la toma de decisiones. En este tipo de labor, es necesaria la generación de modelos que hagan explícita la interacción entre los ecosistemas y la sociedad, y que permitan la visualización simulada computacionalmente de las consecuencias de esta interacción y de las decisiones de los actores involucrados. Por lo que resulta de enorme ayuda contar con indicadores del estado de los ecosistemas (como los suelos) que puedan incorporarse como aproximaciones realistas a estos modelos.
Entonces, ¿cómo hacemos para que la Biodiversidad de Microorganismos del Suelo se considere en la toma de decisiones?
Actualmente, casi todo el mundo está de acuerdo en que las comunidades microbianas tienen una gran influencia en el “buen funcionamiento” de los ecosistemas, incluidos los agroecosistemas o sistemas productivos. Lo que a veces no es tan obvio, es que las decisiones relacionadas con el manejo de los ecosistemas necesitan tomar en consideración a los motores funcionales de estos últimos: los microbios.
En el LANCIS se está desarrollando un proyecto muy ambicioso en el que se caracteriza a nivel socio-ecosistémico distintos cultivos (café, aguacate, maíz, frijol), bajo diferentes condiciones climáticas y manejos agroforestales. Esto significa que se está colectando información sobre la diversidad de microorganismos en el suelo, sobre las características de nutrición y textura del suelo, sobre diversos aspectos del estado nutricional de las plantas, y sobre la diversidad de herbívoros y polinizadores. Al mismo tiempo, se tiene información acerca de las distintas prácticas de aprovechamiento del ecosistema y se estudia las motivaciones que llevan a los productores a tomar las decisiones que determinan el tipo de prácticas que llevan a cabo en sus parcelas.
De este modo se espera encontrar indicadores que reflejen la constitución de la biodiversidad del suelo, y que sean aproximaciones realistas del estado del ecosistema. Finalmente, se espera entender cómo la toma de decisiones y las prácticas de los productores se ven reflejadas en el estado de los ecosistemas, y cómo ellos perciben las consecuencias ecosistémicas de sus intervenciones. Idealmente, dentro de un esquema transdisciplinario, ésta debería ser información producida de manera conjunta, con el potencial para influir en futuras decisiones.
Bibliografía:
Escalante, A. E. (2017). Sistemas productivos: microorganismos, suelos y sostenibilidad. Oikos. Instituto de Ecología UNAM(18), 24-27.
Los hongos comestibles podrían ser el alimento del futuro, ya que poseen una gran cantidad de proteínas fácilmente digeribles. El champiñón, por ejemplo, tiene aproximadamente 80% de agua y el 20% restante es materia seca. De ese 20 %, el 40 % es proteína, además de que contienen grasas, vitaminas y minerales, señala la Maestra en Ciencias Elvira Aguirre Acosta, encargada de la Colección de Hongos del Herbario Nacional de México del Instituto de Biología de la UNAM.
En la época prehispánica los hongos eran llamados nanacatl, por el parecido que tienen en aspecto y sabor con la carne, y eran muy consumidos por los aztecas.
El Herbario Nacional de México es una de las once Colecciones Biológicas Nacionales que tiene bajo su custodia el Instituto de Biología de la UNAM, constituida por colecciones de algas, briofitas, hongos, líquenes y plantas vasculares, incluyendo sus colecciones anexas.
Aquí, nos referiremos específicamente a la Colección de Hongos, iniciada en 1947 por los especialistas Manuel Ruiz-Oronoz y Teófilo Herrera. Actualmente cuenta con más de 30 mil ejemplares que representan alrededor de 400 géneros y mil 600 especies de macromicetos, que son hongos «superiores».
La mayoría de las setas y hongos de «sombrero» son Basidiomicetos. Los Ascomicetos tienen formas diversas, como platos, cazuelas, copas, orejas, u otras formas, cuya característica principal es la presencia de ascosporas (esporas sexuales), encerradas en unos pequeños sacos llamados ascas. En tanto que los Mixomicetos presentan fructificaciones pequeñas de formas muy variadas, que producen esporas con una pared celular. La mayor parte de su ciclo de vida presentan estructuras desnudas, es decir sin pared celular.
Esta colección conserva solamente hongos macroscópicos deshidratados, adaptados para su consulta. Este tipo de hongos poseen un cuerpo formado por un conjunto de filamentos o hifas y en condiciones adecuadas de temperatura y humedad forman cuerpos fructíferos productores de esporas, además, pueden presentar estructuras accesorias como el sombrerillo de las setas. Los hongos microscópicos también pertenecen al reino Fungi, por su pequeño tamaño no se pueden observar a simple vista, se albergan en las colecciones de cultivos, detalla la investigadora.
Los especímenes, dice, proceden de la mayoría de las entidades federativas de la República Mexicana, y por ende, de diversos tipos de ecosistemas. Por esto, la colección tiende a ser representativa de la biota del país, siendo más numerosos los ejemplares de la Ciudad de México y de los estados de Hidalgo, Jalisco, México, Morelos, Puebla, Oaxaca, Sonora y Veracruz, particularmente de Bosques templados y de Selvas altas perennifolias y Selvas bajas caducifolias.
Recolección
Tanto el herbario de la UNAM como el del Instituto Politécnico Nacional, fueron los primeros herbarios nacionales que tuvieron colecciones de hongos, en las que prácticamente están representadas la mayor parte de las entidades federativas del país (desde Baja California hasta Quintana Roo), aunque prevalecen todas las recolectas de los estados del centro y algunas del estado de Veracruz y Oaxaca.
De acuerdo con la especialista, la importancia de la Colección de Hongos, igual que todas las colecciones científicas, radica en que todos los ejemplares que contiene cuentan con una etiqueta de datos muy valiosos que indican el lugar de procedencia, es decir, la entidad federativa, localidad específica, tipo de vegetación, altitud y fecha en que fueron recolectados.
Aunque las especies de hongos no se encuentran extintas, muchas están en peligro debido a su vulnerabilidad, puesto que los hábitats han ido cambiando. Muchas de las especies que se encuentran en peligro de extinción, es porque son explotadas para su comercialización, ya que muchas de ellas son comestibles.
La principal función que estos organismos desempeñan en la Naturaleza, es la de ser descomponedores de la materia orgánica, tanto de origen animal como de origen vegetal En un bosque, por ejemplo, es posible encontrar una gran diversidad de hongos en los troncos caídos, ya que degradan la lignina y la celulosa, elementos que constituyen la madera.
Biología, Biogeografía, Taxonomía
Entre las principales investigaciones que se llevan a cabo con la colección de hongos están las taxonómicas (identificación y clasificación), asi como para conocer la diversidad de especies de hongos que existe en el país. Con algunos ejemplares, se han llevado a cabo estudios de Biología molecular y Biogeografía.
La conservación de los hongos recolectados es muy importante. Así, cuando se incorpora un material a la colección éste debe estar muy bien “curado”, lo que significa que el hongo debe estar en buenas condiciones, es decir, que esté bien deshidratado y maduro. Esto es, que haya formado esporas para que el ejemplar tenga validez dentro de la colección.
Una vez que está dentro de la colección, deben realizarse revisiones periódicas, pues en ocasiones algunas plagas pueden introducirse afectando las muestras resguardadas. Cuando esto sucede, el material se lleva a la congeladora para eliminar los huevecillos o las larvas de artrópodos, a fin de eliminarlos, y mantener los ejemplares en óptimas condiciones. Anexa a la colección de macromicetos (hongos “superiores”), se encuentra la colección de fitopatología, iniciada por la Doctora Martha Zenteno en 1958, que en su mayoría alberga plantas parasitadas por hongos. Cuenta con más de 2 mil ejemplares que representan más de 300 especies de hongos organizados taxonómicamente por grupos de patógenos.
La mayoría de las plantas son sensibles al efecto tóxico del Aluminio, pero hay algunas especies que han adquirido mecanismos de tolerancia.
Existen en las plantas tres mecanismos para excluir o evitar la entrada de Aluminio a sus tejidos. Uno es el aumento del pH en la rizósfera, otro es por la producción de mucílago, y el tercero es por la exudación de ácidos orgánicos (AO) o compuestos químicos, como los fenoles (un ejemplo de fenoles es el Resveratrol que se encuentra en la piel de las uvas y, en algunas variedades, en las semillas), cuyas moléculas atrapan o, mejor dicho, secuestran al Aluminio.
Algunas especies que disminuyen la acidez de la rizósfera aumentando el pH mediante la producción de diferentes tipos de exudados son: el trigo (Triticum aestivum), el arroz (Oryza sativa) y los maíces híbridos (Zea mays), así como la planta mutante y modelo de laboratorio Arabidopsis alr-104 (Arabidopsis thaliana).
Entre las que exudan ácidos orgánicos para secuestrar el Aluminio e impedir que entre a las raíces están el trigo, el maíz y el frijol común (Phaseolusvulgaris), las cuales son ejemplos de plantas que liberan ácido málico y/o cítrico. Ejemplos de plantas que liberan ácido oxálico son la acederilla (Rumex acetosella), la planta de té (Camellia sinensis) y el trigo sarraceno (Fagopyrum esculentum).
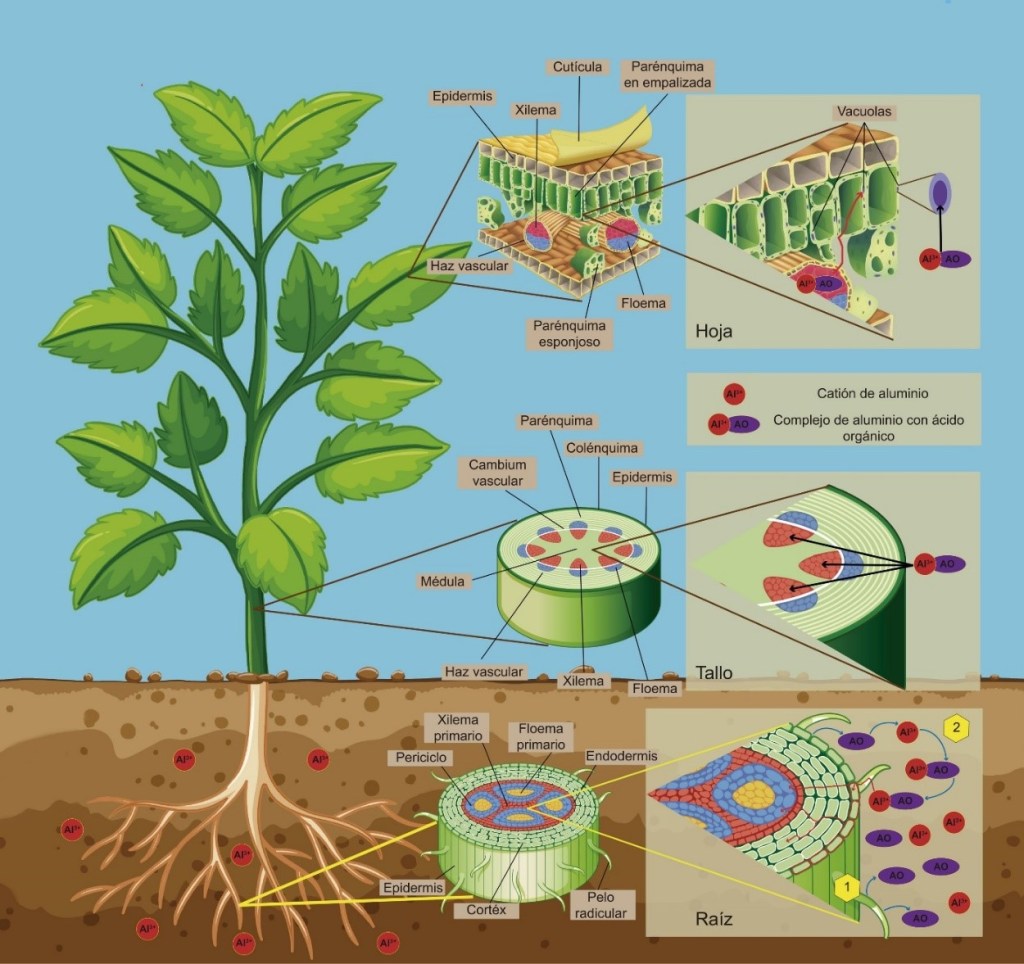
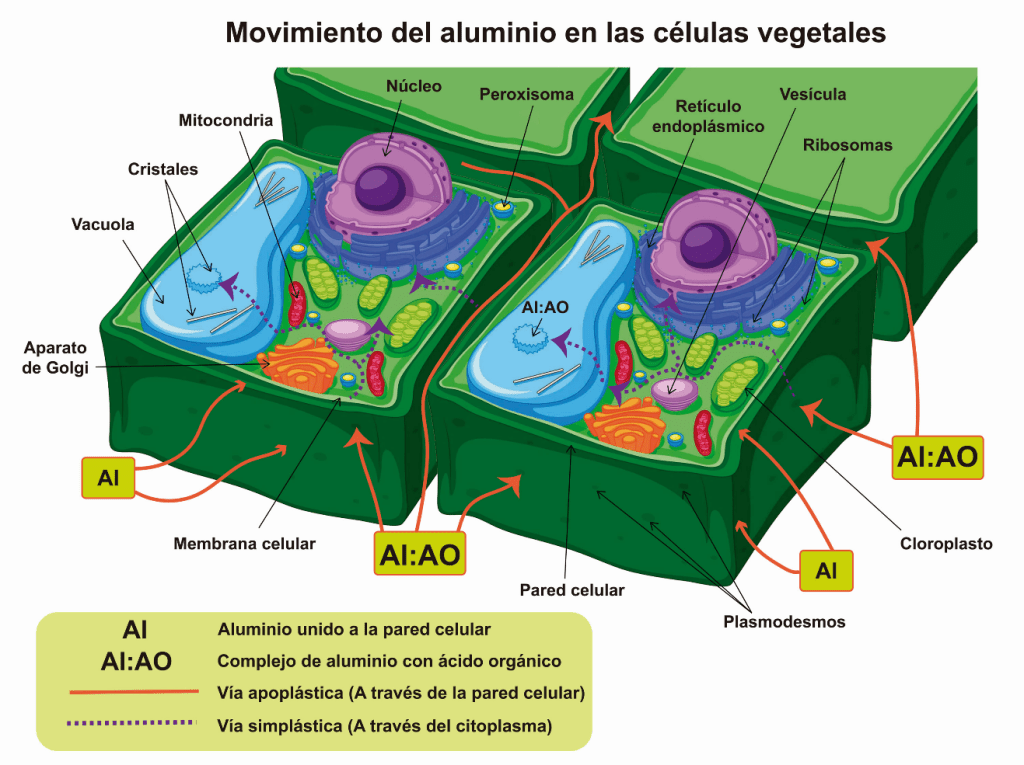
Por otra parte, existe el mecanismo de secuestro del Aluminio, que sucede cuando este metal forma un complejo con un ácido orgánico e ingresa a la raíz, se transporta por el apoplasto o espacio entre las paredes celulares, hasta el simplasto o citoplasma de las células, para almacenarse en las vacuolas. En este organelo el Aluminio continúa formando un complejo con el citrato, malato u oxalato.
Ilustración 1. Acumulación del aluminio en el tejido vegetal
Existen plantas llamadas acumuladoras que pueden aprovechar este mecanismo de secuestro y acumular el Aluminio en las partes aéreas (principalmente en las hojas), es decir, el Aluminio se transporta a través del xilema del tallo hasta las hojas, donde se almacena en las vacuolas. Existen alrededor de 100 especies con estas características dentro de 30 familias, entre las que se encuentran las Asteraceae (familia de la Margarita), Euphorbiaceae (familia de la Nochebuena), Melastomataceae (familia de la Tibouchina), Rubiaceae (familia del café), Polygonaceae (familia del trigo sarraceno) e Hydrangeaceae (familia de las Hortensias).
Un ejemplo fascinante son las hortensias (diversas especies del género Hydrangea), las cuales acumulan grandes concentraciones de Aluminio en las hojas durante los meses de crecimiento y al comenzar la floración, los sépalos se tornan rosas o azules, dependiendo de la concentración de Aluminio en estos tejidos.
Ilustración 2. Relación entre el pH y el aluminio en el cambio de coloración de los sépalos de las hortensias (Hidrangea sp.) Modificado de Schreiber (2014).
Estudios en el Trigo Sarraceno, una especie modelo
En el laboratorio de Alelopatía del Instituto de Ecología, UNAM, se han realizado diversos estudios para conocer los mecanismos de tolerancia del llamado “Trigo” Sarraceno, Fagopyrum esculentum. Esta especie no es un trigo verdadero, es una planta dicotiledónea de la familia Polygonaceae, que se usa, por ejemplo, para hacer crepas, ensaladas o inclusive cerveza.
El Trigo Sarraceno es una planta tolerante y acumuladora de Aluminio, sin embargo, los estudios han demostrado que durante las primeras etapas de desarrollo de la planta (1 a 5 días de edad), es sensible y presenta efectos tóxicos del Aluminio, como la inhibición del crecimiento de la raíz.
Dentro de los estudios del laboratorio de Alelopatía, se puede destacar la investigación en la que se valida la hipótesis de que el Ácido Abscísico (ABA) o fitohormona del estrés, estaba involucrada en la respuesta del Trigo Sarraceno al Aluminio durante sus etapas iniciales de desarrollo. Se demostró que cuando la planta es sometida a concentraciones crecientes de Aluminio, aumentan los niveles de ABA con respecto al tiempo de tratamiento. En este mismo estudio de Ivan Reyna-Llorens y colaboradores, publicaron en 2015 en la revista “Environmental and Experimental Botany”, un posible transportador de Aluminio, nombrado FeALS3, cuyos niveles de transcripción (aumento en la síntesis del ácido ribonucleico o ARN) eran mayores con relación al aumento del ABA. Este estudio mostró que ésta fitohormona (ABA), la cual está involucrada en la respuesta y la adaptación a factores de estrés (como por ejemplo el estrés hídrico o sequía), también puede jugar un papel en inducir los mecanismos de defensa contra el Aluminio, como por ejemplo exudar ácido oxálico en la etapa de plántula.
Recientemente, en un estudio que se publicó en la revista “Plant Physiology and Biochemistry” se demostró que, durante las primeras horas de exposición a Aluminio, se incrementa la actividad de las enzimas antioxidantes como la catalasa, la glutatión reductasa y la ascorbato peroxidasa, las cuales están encargadas de remover a las ERO (especies reactivas de Oxígeno), que como anteriormente se mencionó, son producidas por el efecto tóxico del Aluminio.
Asimismo, Alma Y. Martínez Rendón, como parte de su proyecto de maestría, comprobó que la planta del Trigo Sarraceno expuesta al Aluminio se puede sobreponer a la inhibición radicular de las primeras etapas, y recuperar totalmente su crecimiento durante todo su ciclo de vida, sin mostrar efectos adversos por toxicidad.
Conclusión
Considerando que la acidificación de los suelos es un proceso que avanza rápidamente y que representa una disminución de las tierras cultivables en todo el mundo, las investigaciones relacionadas con el estudio de los mecanismos de defensa, detoxificación y acumulación en las plantas tolerantes, son necesarias para identificar o proponer cultivos alternos que puedan crecer en condiciones de toxicidad por Aluminio sin causar efectos dañinos a la salud humana y al ecosistema.
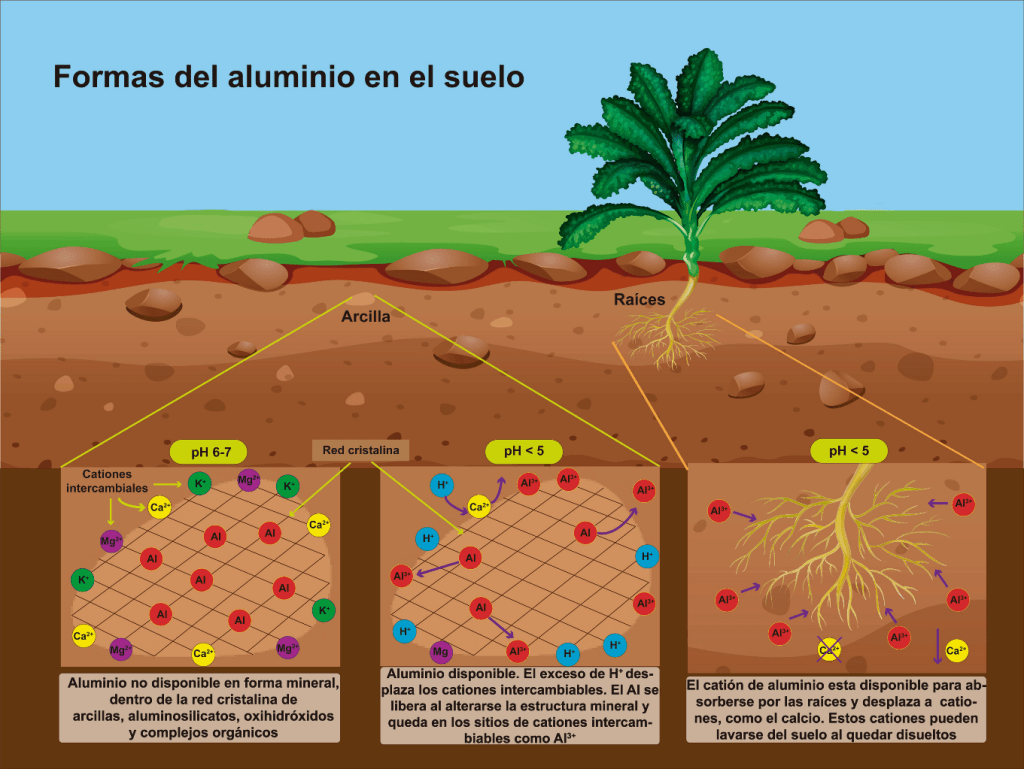
El aluminio (Al) es el tercer elemento químico más abundante en la corteza terrestre. Este elemento generalmente se encuentra unido con otros, por ejemplo, con Silicio, Oxígeno, Calcio, formando diferentes minerales que son parte del suelo. Es importante señalar que el Aluminio es un elemento que reacciona ante los cambios en el pH del suelo. Cuando el valor del pH disminuye (se acidifica), el Aluminio deja de formar parte de los minerales, debido a que la acidez provoca que este elemento se disuelva en la solución del suelo y forme cationes de aluminio (Al3+). Si el Aluminio se encuentra disuelto en el suelo, las plantas lo pueden absorber por sus raíces, y esto es tóxico para la mayoría de las plantas.
Ilustración 1. Formas del Aluminio en el suelo. El Aluminio no disponible, es el que se encuentra dentro de los minerales. El Aluminio intercambiable y soluble, es tóxico para la mayoría de las plantas. Esta figura se obtuvo a partir de imágenes modificadas de brgfx (www.freepik.es)
El Aluminio en los Suelos Acidos
La problemática de los suelos ácidos ha ido en aumento en todo el mundo, principalmente por la creciente demanda en la agricultura que lleva al uso desmedido de fertilizantes que favorecen la acidificación del suelo, esto a su vez origina altas tasas de extracción de nutrientes. En consecuencia, el crecimiento, y la productividad de los cultivos disminuye con el tiempo, lo que representa un grave problema ecológico al aumentar la demanda de suelo para la agricultura.
En el mundo, aproximadamente, el 30% de la superficie agrícola y el 50% de la superficie arable comprende suelos ácidos, y de esta última cifra, el 41% está en nuestro continente. Debido al continuo incremento en los suelos ácidos, y como consecuencia al aumento en la cantidad de cationes de Aluminio (Al3+), que es la forma tóxica de dicho elemento, es importante realizar estudios sobre sus efectos nocivos en las plantas.
Algo importante que ocurre en los suelos ácidos, es que el Aluminio desplaza o sustituye a otros elementos que son indispensables como nutrientes, los cuales al igual que el aluminio en su forma catiónica (Al3+), también tienen carga positiva. Algunos ejemplos son el Calcio (Ca2+), el Magnesio (Mg2+) y el Potasio (K+).
En estas condiciones, el único elemento que pueden absorber las plantas es el Aluminio, y por ello muestran síntomas de deficiencia de nutrientes. Es decir, cuando las plantas absorben Aluminio por las raíces, éste inhibe de manera severa su crecimiento. Pero esto sólo es el reflejo de los cambios que provoca el Aluminio en las funciones metabólicas de la planta, principalmente en la raíz, ya que es el primer órgano que tiene contacto directo con este metal. Además de inhibir el crecimiento longitudinal de la raíz, el Aluminio deforma las raíces, haciéndolas pequeñas y muy gruesas, lo que además disminuye su capacidad de absorber el agua y los pocos nutrientes disueltos en los suelos ácidos.
En la rizosfera, el 95% del Aluminio asociado a las raíces de las plantas se encuentra entre la pared celular y el apoplasto de las células epidérmicas de la raíz. Desde la perspectiva molecular, se ha propuesto que el Aluminio se puede absorber a través de los canales por donde se mueve el Calcio en su forma Ca2+, debido a que ambos elementos son de carga positiva y de diámetro similar; además de que el Aluminio puede desplazar al Calcio de la pared celular y de la membrana plasmática. Cuando el Aluminio sustituye al Calcio, la pared celular se hace rígida y se alteran sus funciones de alargamiento (elongación), que son necesarias para el desarrollo de la célula.
En la membrana plasmática, el Aluminio inhibe la actividad de enzimas tan importantes como la bomba de protones denominada H+-ATPasa. Cómo su nombre lo indica, estas enzimas controlan el flujo de los protones (H+), y por lo tanto alteran el valor del pH afuera y adentro de la célula, así como la absorción activa de los nutrientes responsables del desarrollo celular.
Ilustración 2. Movimiento del Aluminio dentro de las células vegetales. El Aluminio sigue dos vías de transporte: Vía apoplástica (línea anaranjada), en la que se mueve entre las células por la pared celular, y puede ingresar a las células por los plasmodesmos de la pared celular, y Vía simplástica (línea punteada morada) en la que ingresa al citoplasma y se mueve hacia los distintos organelos de la célula. Esta figura se modificó a partir de la imagen de brgfx (www.freepik.es).
Cuando el Aluminio está en el citoplasma de una célula, interfiere con la regulación o concentración de los iones de Hidrógeno (H+), Calcio (Ca2+), Magnesio (Mg2+) y Potasio (K+), provocando alteraciones en el funcionamiento de organelos celulares como las mitocondrias y los cloroplastos. Las alteraciones son principalmente, en la cadena de electrones, lo que da como resultado el aumento en el número de moléculas altamente reactivas, denominadas especies reactivas de oxígeno (ERO), las cuales pueden causar daños irreversibles a las proteínas e incluso al material genético de las células de las plantas.
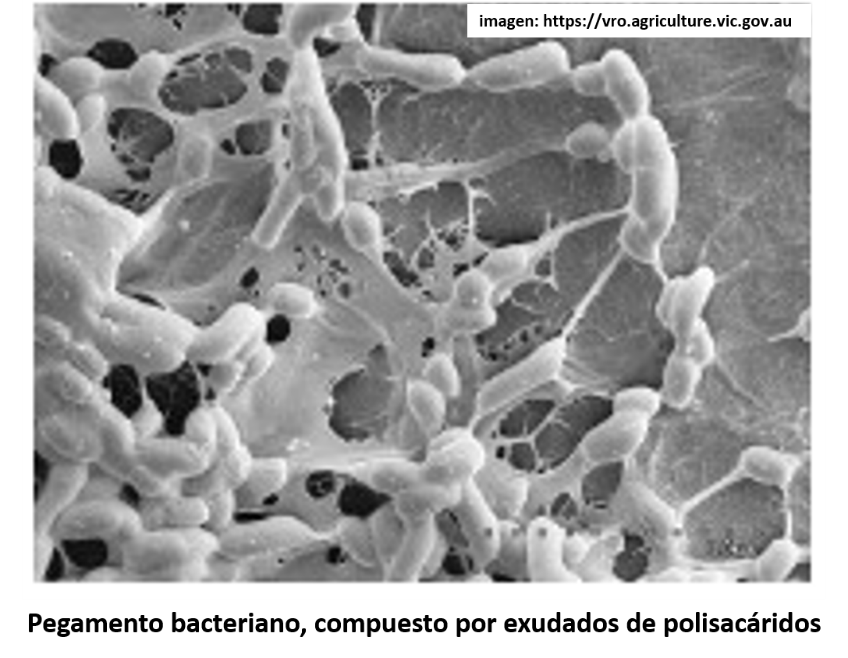
Por otro lado, una respuesta de la raíz ante la presencia de Aluminio es que las células de la epidermis pueden producir un polisacárido o azúcar de cadena larga, conocido como calosa, que se acumula en el espacio intercelular (apoplasto), de las células de las raíces. Esta acumulación de calosa, es un intento de las células vegetales para impedir que el Aluminio ingrese a los tejidos.
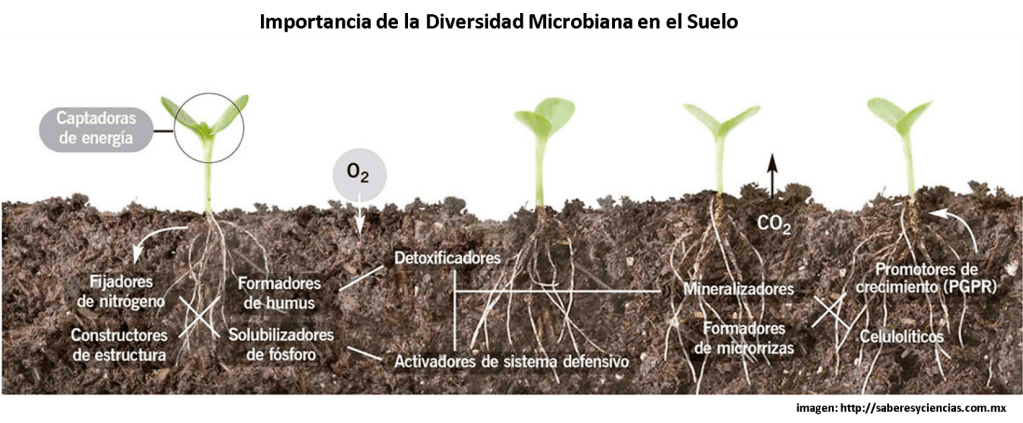
¿Cómo es la vida microscópica en los suelos de los sistemas Tropicales?
El estudio de los microorganismos edáficos en los sistemas tropicales se ha desarrollado muy poco, debido principalmente a la complejidad que este supone. Sin embargo, conocer aspectos ecológicos básicos sobre ellos ha permitido entender su papel en diversos procesos esenciales para mantener el correcto funcionamiento del ecosistema. De manera tradicional se ha abordado el estudio de los microorganismos edáficos en los ecosistemas tropicales clasificándolos según el papel que desempeñan, es decir, en grupos funcionales. Dentro de los organismos más estudiados tenemos a las bacterias fijadoras de Nitrógeno, a los hongos que forman micorrizas, a los organismos fragmentadores de la materia orgánica y a los descomponedores.
Dentro de las principales aportaciones al conocimiento de los microorganismos de suelos tropicales, se han identificado elementos que pueden afectar su abundancia, distribución y permanencia en los ecosistemas.
Se han encontrado patrones definidos en algunos de estos elementos, como pueden ser variables que determinan algunos patrones ambientales:
Temperatura: Algunos organismos principalmente las bacterias, dependen de ella para desarrollar actividades metabólicas, como por ejemplo la respiración. En ecosistemas tropicales el rango óptimo de funcionamiento oscila entre 20 y 24 °C.
Humedad: Es indispensable para el proceso de descomposición y el desarrollo de los hongos. Por el calentamiento global, y el consecuente incremento de la temperatura de en los ecosistemas tropicales, disminuye la humedad, afectando así las funciones que realizan las comunidades de hongos.
Disponibilidad de Nutrientes: Esto es, la facilidad para un organismo de encontrar un recurso. Dependiendo del tipo de nutrientes disponibles podemos encontrar más organismos de un grupo que de otro. Por ejemplo, en suelos en los que el Fósforo es un recurso limitante, los hongos que forman micorrizas serán más abundantes.
pH: Esta variable se mide en un gradiente en dónde los valores más bajos son ácidos y los más altos son básicos. Se ha determinado que es una variable capaz de alterar la dominancia de un grupo sobre otro. Por ejemplo, en suelos ácidos, como los que se encuentran los bosques tropicales, hay más hongos, mientras que en suelos con pH básico (alcalino) dominan las comunidades de bacterias.
A pesar de que en los últimos años se ha hecho un gran esfuerzo por entender cómo se comportan todas estas comunidades microbianas del suelo de distintos ecosistemas, el desarrollo de nuevas tecnologías que sean capaces de facilitar el trabajo de campo para estudiar estos grupos resulta de vital importancia.
Recientemente se han incorporado técnicas que permiten analizar el material genético, mismas que ayudan a conocer la identidad de los organismos, así como sus posibles funciones en el ecosistema. Además, se ha comenzado a experimentar para evaluar la respuesta de estas comunidades ante el cambio de las condiciones ambientales.
Estos ejercicios experimentales servirán para generar información útil para desarrollar alternativas, por ejemplo, la introducción de especies que realicen funciones similares ante escenarios desfavorables (como el del calentamiento global), que podrían provocar cambios en las funciones que los organismos desempeñan de manera natural. Los principales resultados de estos ejercicios sugieren que el aumento en la temperatura y en la concentración de elementos como el Nitrógeno, son capaces de alterar la composición de especies en el suelo de los ecosistemas tropicales, lo que también podría afectar cómo se desarrollan sus funciones.
Es importante señalar que la actividad de las comunidades de microorganismos edáficos influye directamente en el desarrollo y actividad de otros grupos de la biota. Por lo tanto, en un punto de referencia inicial, tener información sobre la diversidad taxonómica y genética de los grupos de microorganismos resulta fundamental para entender la composición y funcionamiento de los grupos que se les asocian, información que no ha sido ampliamente explorada en la actualidad.
¿Qué podemos hacer?
Las fuerzas de presión que se ejercen transformando directa e indirectamente a las selvas, están provocando su deterioro y la pérdida acelerada de estos ecosistemas, de sus recursos y de las especies que en ellos se desarrollan. En México, según el Instituto Nacional de Estadística y Geografía (INEGI), estos ecosistemas abarcan actualmente una superficie total de aproximadamente 4.6% del territorio nacional y han sido ampliamente estudiados en términos de organismos como plantas y animales. Sin embargo, aún son muy escasos los estudios que se han enfocado en conocer la gran diversidad y función que desarrollan los microorganismos del suelo de esos ecosistemas.
¿Hacia dónde vamos? Es necesario establecer prioridades para desarrollar de manera más eficiente las técnicas de investigación que nos permiten entender a las comunidades de microorganismos en suelos tropicales. Pero también se necesita fortalecer la poca investigación que hay, usando herramientas que incorporen modelos matemáticos que nos ayuden a entender los posibles escenarios y que ilustren cómo cambian las dinámicas de funcionamiento de las comunidades de microorganismos en respuesta a las condiciones de cambio a las que se enfrentan, lo que, a largo plazo, se verá reflejado en los procesos en los que están involucrados en los ecosistemas, entre ellos los ciclos biogeoquímicos. Las alteraciones en los ambientes de los microorganismos del suelo pueden tener un impacto global.
En México se conoce poco sobre la gran diversidad de microorganismos que habitan en el suelo, sin embargo, se están haciendo nuevos esfuerzos en el ámbito molecular para conocer su identidad y funciones en los ecosistemas. Además, existe un creciente interés en el estudio de las interacciones que estás comunidades tienen con otros organismos.
La diversidad biológica de la Tierra es el resultado de grandes procesos evolutivos que llegan a durar hasta miles de millones de años. La historia evolutiva de los microorganismos de nuestro planeta es una de las más antiguas. Las comunidades de seres vivos son muy diferentes en distintas partes del planeta debido a esos procesos ecológicos y evolutivos. Por ejemplo, ecosistemas como los desiertos albergan una diversidad de vida menor a la de una selva y entre selvas distintas hay diferencias que podrían ser perceptibles sólo para el ojo experto. Entre las muchas las ideas y preguntas que surgen de la mente de las ecólogas y los ecólogos, algunas buscan entender cómo y por qué las comunidades de seres vivos pueden ser diferentes entre sí, por qué la diversidad es mayor en algunos sitios que en otros y cómo la diversidad diferencial afecta a los procesos ecológicos. En este artículo voy a abordar este tema desde la perspectiva de los ecosistemas tropicales.
Las selvas húmedas, también conocidas como bosques tropicales o selvas altas perennifolias, son consideradas de los biomas terrestres más diversos. Se caracterizan principalmente por su gran complejidad y variedad de nichos ecológicos, que permiten a su vez una enorme riqueza de formas de vida. Está gran complejidad y variedad está relacionada con la manera en la que las especies se desarrollan e interactúan entre ellas en el ecosistema.
Este bioma se encuentra bajo amenaza constante debido a la explotación no sustentable de sus recursos y a presiones de tipo antropogénico; un ejemplo es la deforestación para actividades agrícolas, ganaderas y las diversas actividades relacionadas con la infraestructura urbana. En La evaluación de los cambios de cobertura/uso del suelo en la República Mexicana de Jean-François Mas y colaboradores, se estima que se ha reducido la cobertura de selvas húmedas a un ritmo del 0.76% anual.
En el estudio “Biodiversity recovery of Neotropical secondary forests” de Danaë M. A. Rozendaal y 85 expertos de todo el mundo, se determinó que la recuperación tras un disturbio de mucha intensidad como la deforestación de estos lugares es un proceso largo y complejo. En el artículo, publicado en “Science Advances”, estiman que para recuperar al 80% de las especies eliminadas de estos ambientes al menos se necesita que transcurran 20 años en un proceso de sucesión, es decir que la vegetación se recupere de manera natural, sin intervención humana. Sin embargo, para recuperar la composición inicial, es decir, para volver a tener las mimas especies que existían en el ecosistema antes de la perturbación, son necesarios más de 600 años.
Estos datos son estremecedores y, aunque aparentemente estos ecosistemas tienen una alta capacidad de resistir y recuperarse, el impacto real que las actividades antropogénicas tienen sobre ellos abarca cientos de años.
¿Por qué es necesario conservar los ecosistemas tropicales?
Además de su gran diversidad, ¿por qué son tan especiales los ecosistemas tropicales?, ¿por qué conservarlos? En estos ecosistemas sucede una gran cantidad de procesos importantes que mantienen principalmente el ciclaje de materia y el flujo de energía de nuestro planeta, lo que se conoce como ciclos biogeoquímicos. Estos ciclos químicos llevan el vocablo “bio” porque en ellos participa activamente la biota del sistema y la palabra “geo” porque son de alcance planetario.
Es importante resaltar que la materia orgánica que contienen los suelos de los ecosistemas tropicales es el segundo almacén más importante del mundo de un elemento fundamental para la vida: el Carbono
Los microorganismos edáficos son aquellos que habitan en el suelo y cuyo tamaño se mide en unidades de micrómetros (1 micrómetro equivale a 0.001 mm y, por lo tanto, son sólo visibles con un microscopio). Incluyen hongos, bacterias, algunas algas, protistas y microartrópodos (por ejemplo, ácaros). Aun cuando éstos últimos no son reconocidos como microorganismos por muchos autores debido a que su tamaño puede ser mayor, son un grupo muy diverso y algunos de ellos cumplen con estas características de tamaño. Además, tienen una importante participación en diversos procesos, como la fragmentación de la materia orgánica y las interacciones que establecen con otros microorganismos edáficos, especialmente con hongos y bacterias.
Todos los microorganismos edáficos son reconocidos principalmente por el papel que juegan en el ciclaje de nutrientes como el Carbono (C), el Nitrógeno (N), el Fósforo (P) y el Potasio (K), entre otros. Uno de los ciclos más conocidos es el del Carbono, en el cual hongos y bacterias principalmente, participan activamente en la descomposición de la materia orgánica y en su incorporación en la cadena trófica.
Por otra parte, algunos hongos establecen relaciones simbióticas con las raíces, que permiten el transporte de nutrientes del suelo a las plantas. En las micorrizas, como se le denomina a este tipo de relaciones, los hongos facilitan a las plantas la obtención de recursos como Fósforo y Nitrógeno y, a su vez, las plantas les aportan energía a los hongos en forma de Carbohidratos. Sumado a esto, algunos microorganismos son capaces de modificar la estructura del suelo, y con ello su capacidad de retener nutrientes. Por ejemplo, los microartrópodos edáficos y los hongos al desarrollarse y moverse por el suelo pueden cambiar su porosidad y por lo tanto la capacidad del suelo de retener agua y, en consecuencia, nutrientes.
Si en nuestros sistemas de pastoreo queremos tener un ganado saludable que sea inmune a las enfermedades y los parásitos, requerimos de niveles adecuados de Selenio, Molibdeno y Zinc en el suelo, pero qué pasa si por sus características geológicas no tiene suficiente cantidad de estos minerales en la roca madre. En este caso es posible que se deban hacer aplicaciones de dichos nutrientes una vez cada 3 o cada 5 años, incluso quizá solo una vez cada década. El hecho es que necesitamos hacer este tipo de enmiendas, en las que tenemos que aportar lo que sabemos que falta en el suelo debido a su orígen geológico.
Para realizar un análisis geológico se necesita tomar una muestra una sola vez para cada tipo de suelo, ya que evidentemente no va a cambiar. En contraste, el análisis en base a la Capacidad de Intercambio Catiónico (CIC) del suelo se realiza una vez al año para el caso de futales y hortalizas de alto valor comercial y una vez cada 36 meses para el caso de cultivos extensivos.
Hemos estado haciendo el Analísis de Haney durante los últimos dos años. Todavía estamos adquiriendo experiencia en su utilización, es decir estamos desarrollando más datos, sin embargo, lo que se ha vuelto evidente en este tiempo es que el Análisis de Haney* se correlaciona mucho mas considerablemente con la absorción real de nutrientes por parte de las plantas que se reporta en el análisis de savia, que lo que se reporta en el análisis de suelo basado en la CIC. Asimismo, se correlaciona con la respuesta del cultivo, mucho más de lo que lo hace el análisis de suelo. De ahí que, si se adopta una perspectiva diferente, en base a un enfoque diferente adicional, las recomendaciones agronómicas, las recomendaciones de productos y las recomendaciones de fertilización cambian.
Aunque todo depende del contexto, ya que cada granja y cada campo es único, generalmente la forma en que hemos visto que nuestras recomendaciones agronómicas cambian a medida que hemos utilizado el análisis de savia y hemos cambiado nuestra perspectiva respecto al análisis de suelo basado en la CIC, es que normalmente vemos que las aplicaciones de Potasio bajan hasta en un 70 % o incluso más, y que las aplicaciones de Nitrógeno se reducen hasta en un 60 a 70 %; y casi sistemáticamente estamos realizando aplicaciones foliares de Manganeso o Fierro y también es muy común que apliquemos Cobre, Cobalto, Molibdeno y Boro. Esto es lo que usualmente vemos en la mayoría de los entornos agrícolas cuando comenzamos a trabajar con diferentes productores, en parte es el resultado de cambiar nuestra perspectiva sobre lo que está sucediendo con la química del suelo mediante el uso de diferentes tipos de análisis químico, pero también es el resultado de enfatizar nuestro enfoque en la Biología, porque cuando adicionas y fomentas la Biología del suelo, ésta puede proporcionar una gran cantidad de Nitrógeno y Potasa a un cultivo (si esa potasa esta presente en la roca madre).
Debemos preguntarnos qué ha cambiado, ¿qué es diferente hoy en día, de cuando William Albrecht desarrolló su análisis del suelo? (que era la ciencia emergente de actualidad en los años 40´s y 50´s), y ¿por qué lo hizo?
Carey Reams y William Albrecht solían reportar cómo se podían hacer enmiendas de suelo con un par de Toneladas por acre de Cal (5.0 Ton/ha aproximadamente), una tonelada o dos de Fosfato de roca (2.5 a 5.0 Ton/ha aproximadamente) y una Tonelada o dos (2.5 a 5.0 Ton/ha aproximadamente) de estiércol de aves de corral, y ver esas increíbles recuperaciones en las que los rendimientos de Alfalfa y otros forrajes aumentarían entre un 50 y 60 % por ciento en un solo año, e informaron de éstas increíbles respuestas de cultivos no solo una o dos veces. De hecho, si revisamos la literatura histórica y leemos todos los artículos originales de William Albrecht, encontraremos muchos resultados verdaderamente interesantes que se reportaron acerca del trabajo que estaban haciendo con el balanceo del suelo, sin embargo, hoy en dia no los vemos. Rara vez he observado el tipo de resultados y mejoras que William albrecht reportó, sin duda los he visto ocasionalmente; a veces se dan, pero no constantemente. De ahí que he llegado a la conclusión de que ellos pudieron ver esos tremendos cambios porque tenían una Biología del suelo realmente activa. Mi teoría es que, en los últimos 70 años, a partir de que se realizó esa importante investigación, la Biología de los suelos se ha degradado mucho en todas partes, debido al uso generalizado de pesticidas en el medio ambiente.
Además del uso masivo de pesticidas y la degradación de la biología del suelo, actualmente estamos usandoequipos agrícolas mucho más grandes y en consecuencia tenemos una compactación mucho mayor que la que se tenía hace 70 años. La combinación de estos factoresy probablemente también de otros, realmente ha cambiado la forma en que responden nuestros suelos. En otras palabras, creo que todo se reduce a que hoy tenemos suelos relativamente degradados y que no tienen la actividad biológica que estaba presente hace 70 años.
Ahora bien, si sus cultivos producen altos rendimientos con una calidad excepcionalmente alta y están completamente libres de plagas y enfermedades ¿realmente le preocupa si su reporte de análisis de suelo está idealmente equilibrado? ¿o si está en el rango perfecto?
Realmente nos interesa producir cultivos sanos de alta calidad y con altos rendimientos, e históricamente nos han dicho qué si tenemos nuestros suelos óptimamente balanceados desde la perspectiva de la química, lo lograremos. En mi opinión eso no es del todo cierto, ya que aun si tenemos un suelo químicamente equilibrado ideal, todavía debemos tener una biología diversa y activa.
¿Cuál es el objetivo?, ¿tener un rendimiento óptimo del cultivo? o ¿tener reporte de suelo perfectamente balanceado?Son dos objetivos muy diferentes, con una buena biología, podemos tener un rendimiento óptimo del cultivo aun cuando no se tenga un reporte de suelo perfectamente balanceado.
Así que, revise los reportes del laboratorio y observe las plantas, y si ambos no coinciden, será necesario encontrar un mejor análisis de laboratorio, porque en el que está confiando probablemente no le esté brindando un indicador preciso de lo que está sucediendo. Si es posible, opte por usar laboratorios analíticos cuyos datos se correlacionan con la observación de campo 10 veces mejor que los del análisis de suelo tradicional. Estos son un indiscutible complemento para el análisis de savia porque se correlacionan muy claramente con la observación de lo que ocurre en campo.
Fuente: “Soil Testing 2.0: Build your Soil, Cut Costs & Increase Profits with Regenerative Agriculture”
Cuando pensamos en el manejo del suelo, hay que contemplar 4 aspectos diferentes. Además de la estructura, la química y la biología que mencionamos anteriormente, también está la geología del suelo.
Evidentemente hay pocas cosas que podemos hacer para cambiar la geología del suelo, pero podemos equilibrarla, preservarla y encontrar formas de hacer sinergia con ella. De ahí que, considero que debemos abordar la nutrición vegetal desde una perspectiva biológica, no desde una perspectiva química, y a partir del marco de las ciencias emergentes que estamos desarrollando y aprendiendo del Dr. James White y otros, acerca de cómo la biología alimenta a las plantas; deberíamos tratar de obtener toda la nutrición vegetal a partir de la biología y no de la química.
Dentro de ese contexto, el uso de análisis de suelos basados en la química realmente nos ha dado un par de anteojeras, es decir, ha cegado nuestra perspectiva al centrarnos solo en ese aspecto, por eso, si queremos manejar bien el suelo, debemos tratar de utilizar análisis que describan su biología, estructura y geología, y en todo caso los resultados de estos análisis deben tener tanto peso para nosotros como el del análisisquímico.
Generalmente hemos encontrado que los únicos nutrientes del análisis de suelo que tienden a correlacionarse con el análisis de savia, son el Azufre, Zinc y Boro. Esta es una lista muy corta, tan solo 3 nutrientes de un total de 16 ó tal vez 12. De hecho, en un típico análisis de suelo se reportan 12 nutrientes, por lo que 3 nutrientes de 12 es el 25%.
Entonces, ¿cómo usamos el análisis de suelo hoy en día? Los productores con los que trabajamos todavía realizan el análisis del suelo con base a la CIC. Este tipo de análisis es bien conocido y tiene cierta utilidad, aunque limitada.
La manera en la que nosotros lo usamos es específicamente para manejar el equilibrio Calcio/Magnesio de nuestro suelo (variable en función de la geología), ya que encontramos que este tiende a ser muy valioso e importante. También los usamos para evaluar el Azufre, Zinc y Boro, y eso es todo. Todo lo demás depende del contexto, y ese contexto es la geología, la biología y la estructura del suelo, que son los otros 3 aspectos que deberíamos estar midiendo.
Hemos ampliado nuestro enfoque actual acerca del manejo del equilibrio mineral del suelo y en lugar de solo obtener los datos de éste análisis basado en la CIC, también estamos buscando recopilar información a partir de otros dos tipos de análisis. El primero es un ensayo geológico total, que esencialmente es un análisis de minería, en el que se envían muestras de mineral y muestras de roca a un laboratorio para correr un análisis completo de todo lo que está contenido en ellas. Actualmente hay algunos laboratorios que realizan este tipo de análisis: Laboratorios Acme, en Columbia Británica, Agent Lab en Ontario y Midwest Lab, en el cual también se hacen análisis de suelos y con el que muchos agricultores están familiarizados. En mi opinión, en este último no realizan un análisis geológico completo, pero sí, uno para todos los minerales importantes desde el punto de vista agronómico.
El análisis geológico se realiza solo una vez, no es algo que deba repetirse año tras año, y nos dirá el contenido total de cada nutriente que existe en el suelo, por ello la perspectiva que nos da es muy valiosa.
Hace un par de meses, David Miller, uno de nuestros consultores, estaba revisando un análisis del suelo del Medio Oeste
(no recuerdo si era de Kansas o Nebraska) y un análisis de suelos de Florida. El análisis de CIC también contenía los resultados de los ensayos geológicos, y si no mal recuerdo ambos tipos de análisis se realizaron en los laboratorios Midwest. En otras palabras, tenia los datos del análisis geológico y quimico de todas las muestras que se extrajeron al mismo tiempo en ambos sitios.
Ahora bien, tanto para el suelo de Florida como para el del Medio Oeste, el análisis de CIC reportó 10 partes por millón (ppm) de Manganeso.
Por su parte, el análisis geológico, del suelo de Florida también mostró 10 ppm de Manganeso, lo que significa que el 100% de lo que había en el suelo, aparecía en el análisis de CIC, pero para el suelo del Medio Oeste se reportaron 400 ppm de Manganeso. Por consiguiente, si en el suelo de Florida queremos tener niveles de Manganeso superiores a 10 ppm en su solución (que fue lo que evaluó el análisis de CIC), es obvio que vamos a tener que adicionarlo porque no está presente en esos suelos arenosos, sin embargo, para el caso del Medio Oeste es todo lo contrario. Es decir, si en el suelo del Medio Oeste ya tenemos 400 ppm de Manganeso en las primeras 6.0 pulgadas (15.0 cm aproximadamente), no necesitamos aportar más cantudad de este elemento, simplemente necesitamos darle a la biología del suelo las fuentes de alimentos que necesita para aprovecharlo, esto es, extraerlo y ponerlo a disposición del cultivo,
Esta información nos da una perspectiva muy valiosa sobre cómo debemos manejar las enmiendas del suelo, lo cual es fundamental en el contexto de la agricultura regenerativa.
Ya existe la conciencia en algunos productores, consultores y agrónomos de que nuestro objetivo debe ser desarrollar un ecosistema agrícola que sea tan robusto y saludable que no necesitemos estar aportando insumos constantemente. Debemos eliminar la necesidad de insumos, no obstante, es posible que necesitemos hacer enmiendas ocasionales para suplementar los minerales que debido a la geología de nuestro suelo no están presentes en el perfil, ya que ésta es una condición muy importante.
Fuente: “Soil Testing 2.0: Build your Soil, Cut Costs & Increase Profits with Regenerative Agriculture”